|
|
L'INFORMATION SENSORIELLE EST INTÉGRÉE AU NIVEAU DES VOIES ET DES CENTRES SENSITIFS
|
|
|
1. PRINCIPES D'ORGANISATION DES VOIES SENSORIELLES PRIMAIRES
|
| Chaque voie comporte une succession de plusieurs neurones d'ordre 1, 2, 3 - le neurone d'ordre le plus élevé ayant comme destinée finale l'aire corticale réceptrice de projection correspondante à la modalité sensorielle stimulée (cortex auditif, somesthésique ). Le neurone de premier ordre a son corps cellulaire soit à l'intérieur du système nerveux central (vision, olfaction) soit dans un ganglion périphérique (audition, vestibule, somesthésie, gustation). Le prolongement périphérique de ce neurone (dendrite) est soit une terminaison libre soit innerve une structure plus ou moins complexe constituant un récepteur spécialisé formé à partir d'éléments cellulaires variés (cellules épithéliales ). À l'exception de la voie olfactive, dont la terminaison finale se situe au niveau du rhinencéphale, tous les autres systèmes ont une aire de projection primaire sur le cortex après un dernier relais dans un noyau spécifique du thalamus.
Une des caractéristiques fondamentales de l'organisation des voies sensorielles primaires est l'existence d'une correspondance topique, c'est à dire point par point, entre le récepteur périphérique et son aire de projection corticale. Ceci vaut pour la somesthésie et la vision, c'est à dire pour des récepteurs à extension spatiale (surface du corps / rétine) comme pour l'audition (cochlée). À priori, cette organisation topique, point par point, laisse supposer l'existence de chaînes de transmission linéaires du récepteur jusqu'au cortex. Il existe, cependant, des phénomènes de convergence et d'inhibitions latérales entre les chaînes parallèles.
|
|
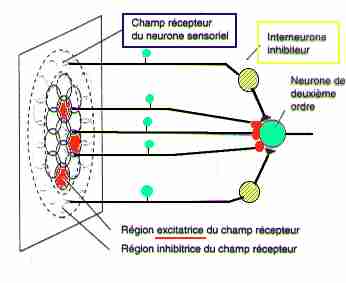
Plusieurs récepteurs peuvent être sous le contrôle d'un même neurone de premier ordre. De même, plusieurs neurones de premier ordre peuvent contrôler un même neurone de second ordre. Dans la rétine, par exemple, les cônes (couleur), localisés au centre de la rétine, disposent d'une chaîne linéaire tandis que les bâtonnets (brillance), disposés à la périphérie, ont une chaîne afférente convergente. L'acuité visuelle est donc bien plus grande au centre de la rétine qu'à la périphérie.
Au niveau périphérique, les terminaisons nerveuses se chevauchent souvent en innervant les récepteurs voisins, ce qui implique une superposition des champs récepteurs. Mais, la discrimination spatiale est permise du fait de l'existence d'interconnexions inhibitrices entre les chaînes parallèles, ce qu'il a été convenu d'appeler des inhibitions latérales. Les inhibitions latérales augmentent, en effet, les effets de contraste et donc, l'acuité tactile ou visuelle.
|
|
2. ORGANISATION DU THALAMUS ET DU CORTEX SENSORIEL
|
| LE THALAMUS
Le noyau ventro-postérieur (VP) reçoit les afférences de toutes les voies de la sensibilité extéroceptive. Il se projette sur le cortex somesthésique primaire pariétal. Les corps genouillés externes (CG Ext) ou latéraux (CGl) sont les centres de relais des voies visuelles. Ils se projettent sur le cortex visuel occipital. Les corps genouillés internes (CG Int) ou médians (CGm) sont les centres de relais des voies auditives. Ils se projettent sur le cortex temporal auditif. Ces noyaux spécifiques sont organisés somatotopiquement comme les voies ascendantes qui s'y projettent et comme les aires corticales sur lesquelles ils se projettent.
Le noyau latéro-dorsal (LD) reçoit ses afférences des noyaux latéro-ventraux sous-jacents et projette ses efférences vers le cortex pariétal associatif. Le pulvinar, grand noyau intégrateur sensitif, reçoit ses afférences des noyaux relais sensitifs sous-jacents (VPL - CG Ext - CG Int) et se projette dans l'aire dite du "schéma corporel" aux confins des cortex pariétaux, temporaux et occipitaux.
|

Coupes frontales du Thalamus
Parties moyenne et postérieure
|
| LE CORTEX
Chaque modalité sensorielle a ses propres aires de projection corticale : somesthésique (pariétales), visuelle (occipitales), auditive (temporales). Les aires
primaires sont organisées de façon somatotopique
et reçoivent les projections des noyaux relais thalamiques
spécifiques.
Il existe au niveau du cortex une organisation verticale "en colonnes" du traitement de l'information : une colonne corticale est un cylindre de cortex (2-3 mm d'épaisseur) d'environ 300 µm de diamètre. A l'intérieur de cette colonne, les neurones sont activés par le même type de récepteurs situés dans une même zone du corps. Il existe un certain degré de traitement de l'information au sein de chaque colonne corticale. Certains neurones du cortex primaire sont capables de détecter la direction du stimulus dans le champ récepteur. Ils détectent également l'intensité du stimulus : l'intensité de la décharge neuronale est fonction de l'intensité de la stimulation du champ récepteur périphérique. La colonne est l'unité
fonctionnelle de base du cortex cérébral.
Le cortex sensitif est donc
constitué par la juxtaposition d'innombrables colonnes
corticales qui constitue la carte somatotopique du corps
("homonculus sensitif"). Cette organisation est due à la présence d'interneurones inhibiteurs GABA (cf. les neuromédiateurs) dont les axones
horizontaux touchent essentiellement la partie basale des cellules
pyramidales. Ce processus inhibiteur est un renforçateur
de contraste. Ces cartes sont, en fait, multiples : 7 "cartes" somesthésiques - 9 "cartes" visuelles - 4 "cartes" auditives.
|
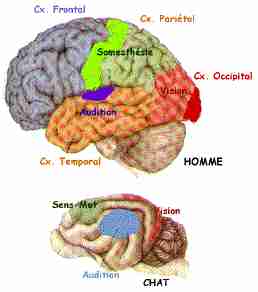
Vue latérale du Cortex Cérébral du Chat et de l'Homme
Noter le développement énorme chez l'Homme des aires associatives ni essentiellement motrices ni essentiellement sensorielles.
|
| Ces cartes corticales sont dynamiques et dépendent de l'information sensorielle reçue. En effet, des modifications des "entrées" sensorielles peuvent entrainer des changements de ces représentations. Ainsi, en interférant avec l'expérience sensorielle - en particulier, au cours du développement - on peut observer des réorganisations importantes des cartes corticales. Cette plasticité neuronale et synaptique - dont tous les processus sont loin d'être connus - si elle joue un rôle majeur au cours du développement - peut être liée à l'apprentissage et à la mémoire. |
| Les aires sensorielles associatives entourent les aires sensorielles primaires. Les informations leur parviennent après relais dans le cortex primaire correspondant et donc après pré traitement de l'information. Elles reçoivent aussi des afférences cortico-corticales provenant d'autres régions du cortex et des afférences thalamiques provenant de certains noyaux non spécifiques du thalamus. Les stimulus auxquels les neurones deviennent les plus sensibles y deviennent de plus en plus complexes. Les aires sensorielles associatives assurent un degré très élaboré de traitement des informations sensorielles. Elles sont, par leurs connexions cortico-corticales, directement impliqués dans de multiples tâches sensori-motrices qui associent, par exemple, les cortex somesthésique et visuel ou les cortex moteur et somesthésique.
Le traitement des messages dans les premiers relais sensoriels est souvent contrôlé par des fibres provenant
des aires corticales. Le rôle de ces fibres corticifuges est variable d'un système sensoriel à l'autre. Il peut être de contrôler directement le fonctionnement des récepteurs sensoriels eux-mêmes.
|
|
