|
|
LE SARCOPLASME CONTIENT LE MATÉRIEL CONTRACTILE DE LA FIBRE MUSCULAIRE
|
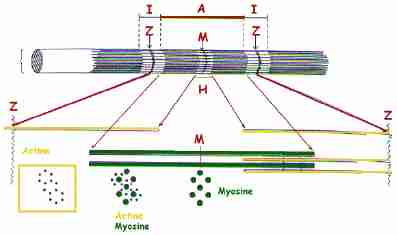
LE SARCOMÈRE : UNITÉ DE CONTRACTION ÉLÉMENTAIRE
|
| Le muscle squelettique est dit "strié" en raison de son aspect au microscope optique. Une bande sombre (bande A : anisotrope) alterne avec une bande plus claire (bande I : isotrope). Chaque bande I est elle-même coupée transversalement par une ligne étroite et sombre : la strie Z. Une zone plus claire, la bande H, partage la bande A en son milieu. Cette striation est due à la présence de filaments contractiles d'actine et de myosine, disposés longitudinalement.
Le sarcomère (longueur au repos = 3 µm), segment de myofibrille compris entre deux stries Z successives, est l'unité de contraction élémentaire Il contient des filaments fins d'actine et des filaments épais de myosine. La bande H ne contient que des filaments épais de myosine; les parties terminales de la bande A contient à la fois des filaments épais de myosine et des filaments fins d'actine; la bande I ne contient que des filaments fins d'actine.
|
| Les filaments fins d'actine
Les filaments
d'actine sont attachés par une de leurs extrémités
à la strie Z et se prolongent parallèlement à
l'axe de la myofibrille jusqu'à la partie médiane
du sarcomère, le long de la bande I puis dans la bande
A. Ils sont constitués de deux chaînes (actine F), enroulées en double hélice, d'une
protéine globulaire : l'actine G et de deux autres protéines : la tropomyosine, qui s'étend
dans la gouttière de l'hélice d'actine, et la troponine
(TN), qui se fixe de place en place sur la tropomyosine (environ toutes les 8 molécules d'actine).
- La tropomyosine bloque les sites de liaison de l'actine et de la myosine.
- La troponine est responsable de la régulation cytoplasmique
de la contraction musculaire. Elle est constituée de 3 peptides :
- la troponine T (TN-T) responsable de la fixation de la troponine sur la tropomyosine;
- la troponine I (TN-I) inhibant l'activité ATPasique (énergie cellulaire) de la tête de myosine;
- la troponine C (TN-C) avec son site spécifique liant le calcium (Ca2+). Lorsque la TN-C est saturée en Ca2+, l'effet inhibiteur de la TN-I est levé.
|
| Les filaments épais de myosine
Chaque filament
épais de myosine se compose d'environ 150 à 360
molécules de myosine, assemblées à la manière
d'une torsade. Chaque molécule de myosine possède
une partie céphalique (tête de myosine), scindée
en deux, siège de l'activité ATPasique. Cette partie
céphalique (tête de myosine) s'articule avec une
partie cervicale, la partie céphalique et la partie cervicale
constituant la méromyosine lourde. Cette méromyosine
lourde est réunie à une partie caudale : la méromyosine légère. La mobilité de la partie cervico-céphalique
(méromyosine lourde), à la manière d'une
articulation, permet la fixation réversible de la myosine avec l'actine (formation du complexe actine-myosine) et le glissement des filaments d'actine et de myosine les uns sur les autres.
|
 LES MÉCANISMES CELLULAIRES DE LA CONTRACTION MUSCULAIRE LES MÉCANISMES CELLULAIRES DE LA CONTRACTION MUSCULAIRE
|
Pendant la contraction ou la relaxation musculaire,
la longueur des filaments d'actine et de myosine reste constante.
En revanche, la longueur des sarcomères varie du fait
du glissement des filaments fins d'actine dans le réseau
des filaments épais de myosine : la longueur de la bande A (longueur des filaments de myosine) reste inchangée, tandis que les bandes H et I ont des longueurs variables. Une cascade d'événements très complexes survient entre le moment où le sarcolème - parcouru par un potentiel d'action musculaire - est dépolarisé et le raccourcissement du sarcomère.
- Le potentiel d'action (dépolarisation membranaire), en se propageant le long des tubules transverses du sarcolème, parvient à proximité des citernes terminales des tubules longitudinaux du réticulum endoplasmique (triade). Cette dépolarisation (par une cascade d'événements) permet l'ouverture des canaux Ca2+ contenus dans la membrane des tubules longitudinaux du réticulum endoplasmique. L'ouverture de ces canaux permet le relargage du Ca2+ (7) contenu dans le réticulum endoplasmique dans le sarcoplasme de la fibre musculaire. La concentration intracellulaire en Ca2+ ([Ca2+i]) passe de 0,01 µm/l au repos à 10 µm/l (x 1 000).
- Le Ca2+ intracellulaire se fixe sur le site spécifique de la troponine C (TN-C). Cette fixation modifie la conformation de la molécule de tropomyosine, qui glisse alors dans la profondeur de la gouttière de la chaîne hélicoïdale d'actine, libérant ainsi les sites de fixation spécifiques de la myosine présents sur la molécule d'actine. Les têtes globulaires de myosine se fixent alors sur les sites spécifiques de l'actine. Dans le même temps, la fixation du Ca2+ sur la TN-C permet la levée de l'inhibition exercée par la troponine I (TN-I) sur l'activité ATPasique de la tête de myosine. Cette activité ATPasique permet la scission (hydrolyse) Mg2+ dépendante de l'ATP en ADP et Pi (phosphate inorganique), scission productrice d'énergie. Tout ceci aboutit à la formation d'un complexe Actine-Myosine-ADP-Pi (A-M-ADP-Pi) (1).
- Le Pi (2), dans un premier temps, puis l'ADP (3), dans un second temps, se détachent de ce complexe, ce qui permet une modification de l'angle formé par les têtes de myosine fixées à l'actine (90° => 50° => 45°), et donc un glissement des filaments d'actine sur les filaments de myosine. La traction au niveau des deux extrémités d'un filament épais de myosine s'effectue en sens opposé : un seul cycle de glissement raccourcit le sarcomère de 2 x 8 nm (1%).
- Le complexe actine-myosine (A-M) reste stable ("complexe de rigidité") et seule la présence d'une nouvelle molécule d'ATP (4) permet la rupture de la liaison entre l'actine et la myosine (5), le redressement des têtes de myosine (45° => 90°) (5) et la formation d'un nouveau complexe myosine-ATP (5). Si la [Ca2+i] est suffisamment élevée, le cycle se reproduit (1). Au cours d'une même contraction, le cycle se reproduit plusieurs fois, en fonction de la fréquence des potentiels d'action émis par le motoneurone alpha. Plus le nombre de cycles est grand, plus le raccourcissement est important : une secousse musculaire peut entraîner jusqu'à 50% de raccourcissement du muscle.
- Le mécanisme prend fin quand la [Ca2+i] est inférieure à 1 µmol/l (concentration de repos) et que les sites calciques de la TN-C sont libres (6). Les canaux calciques du réticulum endoplasmique se ferment (absence de potentiel d'action musculaire) et le calcium cytoplasmique est transporté activement vers les citernes réticulaires.
|
|