LA SENSIBILITÉ MÉCANIQUE MUSCULAIRE ET ARTICULAIRE
|
1. LES TROIS QUALITÉS DE LA SENSIBILITÉ MÉCANIQUE MUSCULAIRE ET ARTICULAIRE
La proprioception nous permet de connaître la position de notre corps dans l'espace et de nos membres par rapport à notre corps. Nous sommes également capables d'apprécier la résistance contre laquelle nous effectuons un mouvement. De fait, la proprioception correspond à trois qualités : la sensibilité à la position, au mouvement et à la force.
|
- La sensibilité à la position nous informe des angles formés par chacune de nos articulations, et donc de la position relative de nos membres entre eux et par rapport au corps. La précision en est faible et elle présente peu ou pas d'adaptation. Elle peut être grandement améliorée par l'apprentissage (ex : tir à l'arc).
- La sensibilité au mouvement correspond à une sensation de vitesse, de direction et d'amplitude. Les seuils de sensibilité pour ces trois paramètres sont plus faibles dans les articulations proximales (épaule) que dans les articulations distales (main).
- La sensibilité à la force se superpose à la sensibilité à la pression (étirement de la peau et pression exercée par un objet à porter). Il est donc difficile de distinguer l'information en provenance des propriocepteurs de celle provenant des mécanorécepteurs cutanés.
|
2. LES MÉCANORÉCEPTEURS MUSCULAIRES ET ARTICULAIRES
Les récepteurs mis en jeu dans la proprioception sont des mécanorécepteurs localisés dans les muscles, les tendons et les articulations.
|
|
LES MÉCANORÉCEPTEURS MUSCULAIRES : LES FUSEAUX NEURO-MUSCULAIRES
|
| Les fuseaux neuromusculaires sont répartis dans la partie charnue du muscle strié. Ils sont entourés d'une enveloppe fibro-conjonctive, étroite à ses deux extrémités et renflée dans sa partie médiane en une capsule remplie de gel. Cette capsule contient et protège la partie médiane de 4-15 petites fibres musculaires, très particulières, dites intrafusales, disposées dans le corps du muscle parallèlement aux fibres musculaires extrafusales.
Les extrémités de ces fibres intrafusales, situées en dehors de l'enveloppe protectrice du fuseau, s'insèrent sur les cloisons conjonctives intramusculaires. Leurs parties polaires, striées, contiennent des myofibrilles contractiles. Leur partie équatoriale contient l'ensemble des noyaux cellulaires de la fibre.
D'après la disposition de ces noyaux, on distingue les fibres intrafusales à chaîne nucléaire (15 µm de diamètre / 3-5 mm de long) des fibres intrafusales à sac nucléaire (30 µm de diamètre / 6-10 mm de long).
Les fuseaux neuromusculaires assurent le contrôle du tonus musculaire
(stimulus = étirement musculaire). Ils sont à
la base du réflexe myotatique.
|
Innervation sensitive
Les fibres sensitives primaires Ia, qui perdent leur gaine de myéline à proximité du fuseau, se ramifient en autant de branches que de fibres intrafusales. Elles s'enroulent autour de la région équatoriale nucléée de chaque fibre intrafusale. Les fibres sensitives secondaires II, qui perdent également leur gaine de myéline à proximité des fuseaux, s'enroulent presque exclusivement autour des parties juxtaéquatoriales des fibres à chaîne nucléaire.
Innervation motrice
Ces fibres intrafusales à chaîne et à sac reçoivent une innervation motrice spécifique. Les fibres gamma, fusimotrices innervent les parties striées contractiles des fibres intrafusales. Ces fibres fusimotrices, en assurant la contraction des fibres intrafusales, modifient l'excitabilité des fibres sensitives innervant les fuseaux, en déformant mécaniquement le récepteur.
|
D'après Figure 2.29 - Psychophysiologie sensorielle. Neurophysiologie Fonctionnelle II. P. Buser et M. Imbert. Hermann Paris - Collection Méthodes.
|
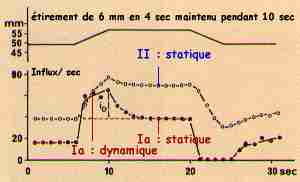
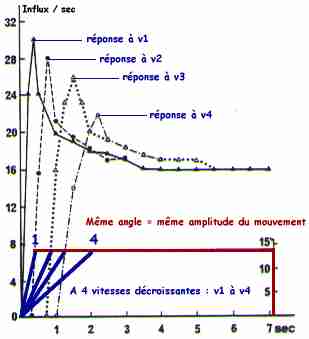
| Les fibres Ia ont (1) une réponse dynamique à l'étirement musculaire, qui dépend linéairement de la vitesse d'étirement du muscle puis (2) une réponse statique pendant tout le temps de l'étirement.
Les fibres II ont essentiellement une réponse statique, avec une réponse dynamique quasiment nulle.
Les fibres primaires (Ia - fibres de 10-12 µm de diamètre) et secondaires (II - fibres de 5-6 µm de diamètre) des fuseaux neuro-musculaires répondent à l'étirement du muscle. Ces récepteurs de longueur ont une adaptation pratiquement nulle : les fibres ont une fréquence de décharge quasi constante pendant tout le temps de l'étirement musculaire. |
| LES MÉCANORÉCEPTEURS TENDINEUX : LES ORGANES TENDINEUX DE GOLGI |
| Les organes tendineux de Golgi sont situés aux jonctions myotendineuses. Ils sont constitués de faisceaux de collagène entourés par une capsule fibro-conjonctive fusiforme. À l'une des extrémités de l'organe tendineux, ces fibres de collagène s'insèrent sur les aponévroses tendineuses. À l'autre extrémité, elles se connectent avec 5-25 fibres musculaires de toute nature appartenant à plusieurs unités motrices différentes. Réciproquement, une unité motrice active 4 à 6 organes tendineux différents par plusieurs de ces fibres constitutives. Il est très important de noter que l'organe tendineux ne s'insère que sur une partie seulement des fibres musculaires : les autres fibres musculaires s'insèrent directement sur le tendon et agissent donc en parallèle avec l'organe tendineux voisin. Leur contraction a tendance à diminuer la déformation de l'organe tendineux, que provoque la contraction des fibres musculaires placées en série sur l'organe tendineux.
Les organes tendineux de Golgi sont innervés par des fibres Ib (fibres de 10-12 µm de diamètre). Ces fibres Ib pénètrent dans la capsule de l'organe tendineux, perdent leur gaine de myéline et se ramifient en plusieurs branches amyéliniques, qui s'insinuent entre les fibres de collagène. Les organes tendineux de Golgi présentent une sensibilité dynamique très développée et renseignent le système nerveux central sur les variations de la force contractile du muscle (stimulus = contraction musculaire active).
|
| LES MÉCANORÉCEPTEURS ARTICULAIRES |
D'après
Figure 2.40 - Psychophysiologie sensorielle. Neurophysiologie Fonctionnelle II. P. Buser et M. Imbert. Hermann Paris - Collection Méthodes.
|
Les fibres myélinisées fines du groupe III et les fibres amyéliniques du groupe IV se terminent par des terminaisons libres.
Les récepteurs de Ruffini des capsules articulaires, les récepteurs de Golgi des ligaments articulaires donnent essentiellement naissance à des fibres myélinisées de gros diamètre (40-60 m. sec-1).
Les récepteurs de Ruffini des capsules articulaires constituent la majorité des récepteurs articulaires. Ils sont à la fois dynamiques et statiques. Ils sont actifs pour un angle précis d'activation, proche des positions extrêmes du membre (flexion ou extension). Lorsqu'un mouvement articulaire est effectué autour de l'angle d'activation du récepteur étudié, on observe une augmentation de la décharge du nerf d'autant plus important que la vitesse de rotation de l'articulation est importante. Lorsque la rotation articulaire est terminée, la décharge nerveuse rejoint le niveau tonique de décharge spécifique de la nouvelle position articulaire.
Ces récepteurs articulaires sont des récepteurs phasico-toniques, qui renseignent sur les mouvements articulaires comme sur la position de l'articulation.
|
